Aktueller Stand des osteogenen Tissue Engineerings

Knochendefekte im Bereich der Kiefer können traumatisch, chirurgisch aber auch atrophisch bedingt sein und stellen weiterhin eine große Herausforderung für die behandelnden Ärzte dar. Durch den zunehmenden Altersdurchschnitt in der Bevölkerung bekommen die Therapie von periodontalen Knochendefekten und die Therapie von Kieferatrophien zur implantatbasierten Rehabilitation des Kauapparates einen zunehmend wichtigen Stellenwert. Allen Krankheitsbildern ist gemein, dass die Wiederherstellung des knöchernen Gesichtsskeletts mit seinen funktionellen und ästhetischen Aspekten für die Lebensqualität der Patienten von großer Bedeutung ist. Die Chirurgie hat in den letzten Jahrzehnten einen Wandel von den eher Resektionsorientierten Verfahren hin zu den auf die Gewebsregeneration fokussierenden Verfahren durchlaufen. Anfangs erfolgte die Skelettrekonstruktionen mittels artifizieller Materialien, wie Metall, Keramiken und Polymeren [Binderman et al., 1990; Frenkel et al., 1975; Leake et al., 1976; Niwa et al., 2000; Pochon et al., 1991]. Somit konnte zwar die Skelettform und teilweise auch die Funktion wiederhergestellt werden, jedoch wurden materialspezifische Nachteile in Kauf genommen. Um langfristige Behandlungserfolge zu verzeichnen, findet mittlerweile zunehmend Berücksichtigung, dass das Skelett einem dynamischen Geschehen unterworfen ist, gekennzeichnet durch einen adaptativen Knochenan- und -abbau. Die Knochenneubildung basiert auf der Fähigkeit vieler ossärer Gewebe, dynamische Umbauprozesse in Gang zu setzten, die im optimalen Fall zu einer Restitutio ad integrum führen, beispielsweise bei der Knochenheilung nach einer Fraktur [Rodan, 1992]. Basierend auf diesen Erkenntnissen kam es zu einer Verwendung von autologen Transplantaten zur Knochenregeneration. Diese besitzen im Gegensatz zu den artifiziellen Materialien lebende, biologisch aktive Zellen, die sowohl an der Einheilung, als auch an der Adaptation an die lokalen mechanischen Anforderungen beteiligt sind. Aufgrund der vielen Vorteile kann hier vom „Gold-Standard“ bezüglich der Knochenrekonstruktion gesprochen werden [Pretorius, 2005]. Neben nicht vaskularisierten, freien Knochentransplantaten werden auch mikrovaskulär anastomosierte Transplantate (hohe mechanische Kompetenz) [Mehta et al., 2004] verwendet, welche bei größeren Defekten oder in vorgeschädigten Geweben Vorteile bieten [Smolka et al., 2005]. Neben der Morbidität aufgrund des Entnahmedefektes [Nkenke et al., 2001; Nkenke et al., 2004; Sasso et al., 2005] ist auch die quantitative Limitation des zur Verfügung stehenden Knochenreservoirs als Nachteil zu nennen. In den letzten Jahren hat insbesondere in der Forschung ein Wandel stattgefunden. Sie zielt zunehmend auf zellbasierte Knochenregenerations- und Rekonstruktionsansätze ab. Dabei unterstützen die vitalen Zellen die Wiederherstellung der Gewebefunktionalität [Langer et al., 1993]. Prinzipiell gibt es drei verschiedene Arten von zellbasierten Knochenregenerationsstrategien. Neben der Transplantation von Zellen in einem Gewebeblock (klassisches Knochentransplantat) ist auch die In-situ-Aktivierung von ortsständigen Zellen (Distraktionsosteogenese) möglich. Hierbei werden körpereigene Zellen durch verschiedene Stimuli, wie Dehnungsreize [Meyer U. et al., 1999a, 1999b], elektromechanische Reize [Aaron et al., 2004] und Stimulation durch Zytokine, zur Knochenbildung angereizt [Kübler et al., 1998; Depprich et al., 2005]. Des Weiteren besteht die Möglichkeit der Transplantation von isolierten und/oder extrakorporal kultivierten Zellen, gegebenenfalls auch als extrakorporal generiertes Gewebekonstrukt. Letzteres wird unter dem Begriff „Tissue Engineering“ verstanden. Hierbei werden lebende Zellen in vitro auf beziehungsweise in einer Matrix kultiviert, um ein Zielgewebe zu erhalten, welches dann in den selben Organismus implantiert werden kann, um spezifische Gewebefunktionen wiederherzustellen. Die drei wichtigen biologischen Säulen des „Tissue Engineering“ sind die Zellen [Handschel et al., 2006], Wachstumsfaktoren [Kübler et al., 1998; Depprich et al., 2005] und die Matrix (Biomaterial) [Handschel et al., 2002; Wiesmann et al., 2004] (Abbildung 1). Durch den Wegfall der Entnahmemorbidität und der Vorteil des unbegrenzten Reservoirs bietet die extrakorporale Herstellung von Ersatzgeweben deutliche Vorteile gegenüber der Verwendung von Transplantaten.
Zellen
Zellarten, die beim „Tissue Engineering“ Verwendung finden, können autologen, allogenen oder xenogenen Ursprungs sein. Diese Kategorien können noch entsprechend ihrer Differenzierungskapazität in ausdifferenzierte (Osteoblasten), multipotente, pluripotente und totipotente Zelllinien unterteilt werden. Im Gegensatz zu totipotenten Zellen, die einen kompletten Organismus generieren können, können sich pluripotente Zelllinien in Zellen aller drei Keimblätter differenzieren. Ihnen fehlt aber die Kompetenz, einen komplexen Gesamtorganismus zu bilden.
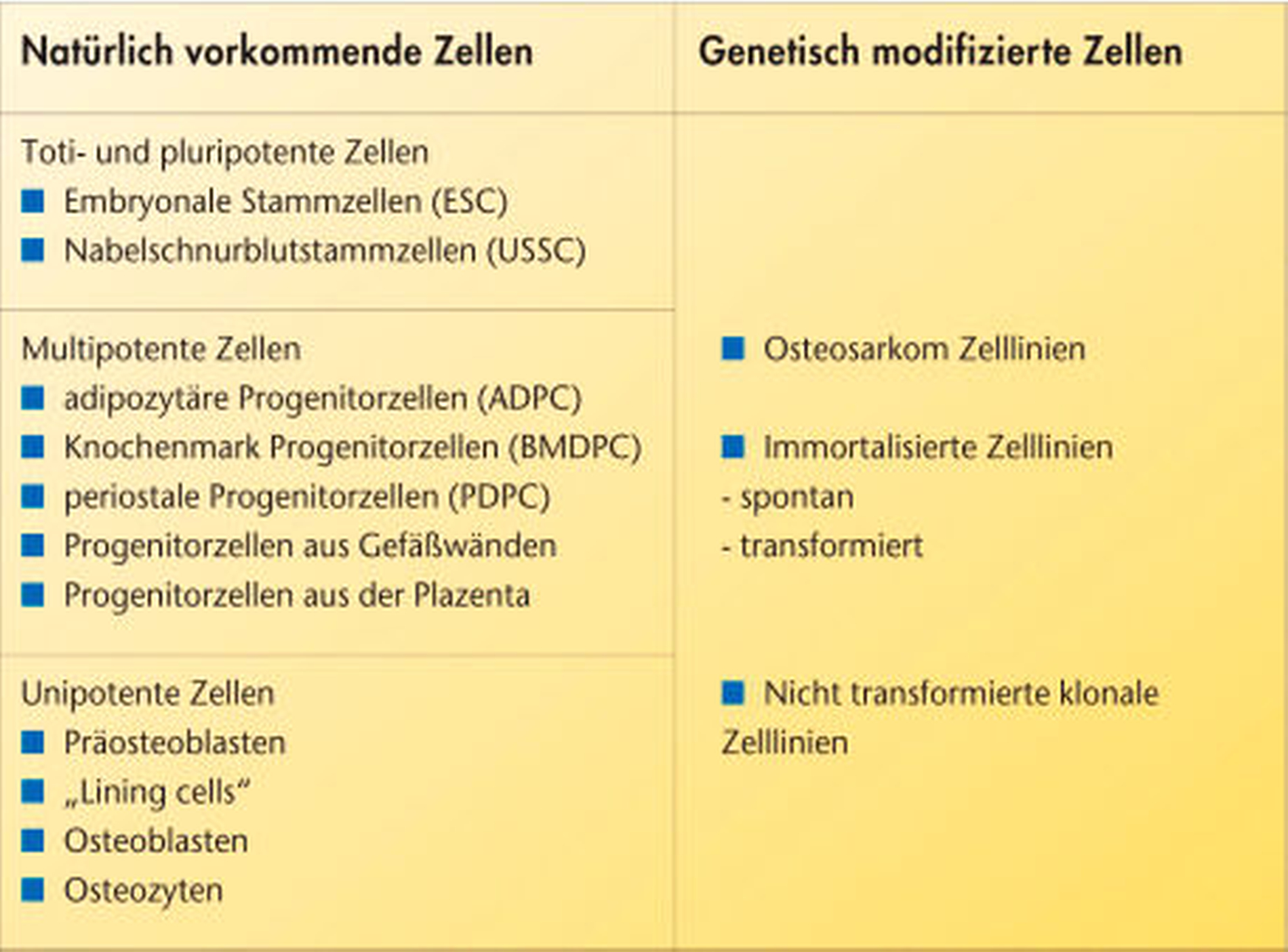
Multipotente Zellen hingegen können in verschiedene Zelltypen differenzieren ohne zugleich pluripotent oder totipotent zu sein. Neben dieser Einteilung der Zellen besteht auch die Möglichkeit, Zellen in natürliche und genetisch modifizierte Zellen einzuteilen (Tabelle, Abbildung 2). Der Vorteil bei der Verwendung von pluri- , toti- beziehungsweise multipotenten Zelllinien (wie pluripotente embryonale Stammzellen (ESC) oder unrestringated somatic stemm cells (USSC)) ist, dass eine Differenzierung der Ausgangszellen nicht nur in die Zellen, die für die Knochenbildung/-mineralisierung verantwortlich sind, erfolgt, sondern auch in Zellen, die weitere wichtige Funktionen bei der Generierung von Geweben übernehmen, zum Beispiel Endothelzellen. Multipotente mesenchymale Progenitorzellen (MPC) stellen eine sehr heterogene Gruppe dar und können aus verschiedenen Geweben (wie Knochenmark, Periost, Fett, und mehr) [Pittenger et al., 1999; Moosmann et al., 2005; Covas et al., 2005; In´t Anker et al., 2004; Wulf et al., 2004; Seo et al., 2004; Ng et al., 2005; Sakaguchi et al., 2004] gewonnen werden und in unterschiedlichen mesenchymale Zelllinien differenziert werden [Handschel et al., 2006]. Stammzellen und Vorläuferzellen entstehen im Embryo und scheinen im adulten Organismus im Gewebe zu persitieren. Dort tragen sie wesentlich zur Reparatur von Gewebedefekten, durch die Differenzierung in Fibroblasten, Osteoblasten, Chondrozyten und Adipozyten, bei [Pittenger et al., 1999; Caplan, 2001]. Die Reifung dieser Zellen inklusive Proliferation und Differenzierung erfolgt unter dem Einfluss verschiedenster Hormone und Zytokine. Die Differenzierung in einen Osteoblasten ist definitionsgemäß dadurch erreicht, dass die Zelle Knochenmatrix synthetisieren kann, die mineralisiert wird. Des Weiteren exprimieren Osteoblasten phenotypische Marker, wie Alkalische Phosphatase, Kollagen Typ I und Osteocalzin [Aubin et al., 1996]. Die Entnahmemorbidität ist verglichen mit einer Knochentransplantatentnahme zwar deutlich geringer, jedoch muss auch bei dieser Zellart eine Entnahme vor der Differenzierung erfolgen, da sie stets autologen Ursprungs sind. Da die MPC in Knochenmarksaspiraten eine gering repräsentierte Subpopulation darstellen – nur eine von 100 000 Zellen aus dem Knochenmark ist eine MPC [D´Ippolito et al., 1999; Quarto et al., 1995] – ist die Gewinnung dieser Zellen recht aufwendig. Ein weiterer Nachteil dieser Zellen ist das begrenzte Erneuerungspotenzial verglichen mit zum Beispiel ESC und USSC [McCulloch et al., 1991]. Erschwerend kommt hinzu, dass diese Zellen in höherem Lebensalter ein vermindertes Proliferationspotenzial und Differenzierungskapazität besitzen [D´Ippolito et al., 1999; McCulloch et al., 1991; Quarto et al., 1995].
Zu den pluripotenten Zellen gehören die embryonalen Stammzellen (ESC), welche aus der inneren Zellmasse der Blastozyste gewonnen werden und eine pluripotente embryonale Vorläuferzelle repräsentieren. Vor über 20 Jahren gelang es erstmals, ESC aus der Maus zu isolieren und zu kultivieren [Martin, 1981]. Ende des letzten Jahrtausends gelang schließlich auch die Etablierung einer stabilen humanan ESC-Zellinie [Thomson et al., 1998]. ESC konnten früher nur in Co-Kulturen mit mitotisch inaktiven Fibroblasten, die als sogenannte Feedercells fungierten, kultiviert werden [Bielby et al., 2004; Heng et al., 2004; zur Nieden et al., 2003]. Dies ist aber unter Anwendung spezieller Faktoren nun nicht mehr notwendig. Unter speziellen Kultivierungsbedingungen können diese ESC in osteoblastenähnliche Zellen (OLC) differenzieren. Dies wird meistens durch den Zusatz von Dexamethason, Ascorbinsäure und ß-Glycerophosphat (DAG) zum Nährmedium erreicht [Bielby et al., 2004; Chaudhry et al., 2004]. Aber auch Zytokine, wie BMP-2, oder Vitamin D3 können die osteoblastenähnliche Differenzierung fördern [zur Neiden et al., 2003]. Anhand von murinen ESCs, die in Anwesenheit von LIF (leukemia inhibitory factor), DAG (Dexamethason, Ascorbinsäuren, ß-Glycerophosphat) oder BMP-2 kultiviert wurden, konnte unsere Arbeitsgruppe zeigen, dass die Kultivierung mit DAG eine bessere osteogene Differenzierung zur Folge hat als mit BMP-2. Dies wurde mittels immunhistologischer Färbungen und einer real-time PCR Expressionsanalyse osteogener Marker (Kollagen I, Osteocalzin) nachgewiesen [Handschel et al., 2008a]. Ein Vorteil der ESCs gegenüber den multipotenten mesenchymalen Progenitorzellen ist, dass sie als immortal bezeichnet werden und ein unbegrenztes Reservoir an differenzierten Osteoblasten für Transplantationen darstellen. Die immunogene Abstoßung dieser Zellen durch den immunkompeten-
ten Transplantatempfänger zu vermeiden, ist eine der wesentlichen Herausforderungen der heutigen Forschung. Interessanterweise konnten Burt et al. eine Major Histokompatibilitäts Komplex (MHC)-mismatched ESC-Transplantation bei Mäusen ohne den klinischen oder histologischen Nachweis einer graft-versus-host Reaktion durchführen [Burt et al., 2004]. Des Weiteren beschrieb Zavazava in einem Übersichtsartikel die Möglichkeit, dass ESC ein Potenzial zur Induktion einer Immuntolleranz besitzen [Zavazava, 2003]. Neben all diesen Vorteilen gibt es auch Bedenken bezüglich der Anwendung der ESC für die Transplantation. So ist eine mögliche maligne Entartung der Zellen denkbar. Trounson beschrieb Beispielsweise das vermehrte Auftreten von Teratomen und Teratolarzinomen bei Tieren mit transplantierten undifferenzierten ESC [Trounson, 2002]. Andererseits haben viele Autoren in ihren jeweiligen In-vivo-Untersuchungen mit ESC keinerlei Hinweise auf tumoröse Entartungen finden können [Zhang et al., 2001]. Zuletzt bleiben die rechtlichen und ethischen Bedenken für die Verwendung von humanen ESC in wissenschaftlichen Versuchen beziehungsweise in der klinischen Anwendung. Die Diskussion bezieht sich letztendlich auf die Frage, ob der Schutz und die Integrität des humanen Embryos oder aber potenzielle Therapieoptionen für Patienten ein höheres Gut darstellen. Verbunden damit ist die Diskussion, ob der absolute Respekt vor dem individuellen menschlichen Leben bereits mit der Konzeption oder erst später beginnt [Cogle et al., 2003; Gilbert, 2004]. Eine mögliche Alternative stellen die vor wenigen Jahren entdeckten mesenchymalen Stammzellen aus dem Nabelschnurblut, sogenannte Unrestringated somatic stem cells (USSC), dar [Kogler et al., 2004]. Bei diesen Zellen handelt es sich um mittels Dichtegradientenzentrifugation gewonnene pluripotente, mononukleäre Vorläuferzellen aus dem Nabelschnurblut mit nur einem geringen immunogenen Potenzial. Das geringe immunogene Potenzial wird schon seit Jahren bei der Behandlung von Leukämien in der Hämatoonkologie genutzt, wo trotz heterologer Transplantation nicht HLA-identischer Individuen wenige Abstoßungsreaktionen auftreten [Benito et al., 2004]. Diese Zellen besitzen neben einem chondroblastären, adipozytären, hämatopetischen und neuronalen Differenzierungspotenzial, auch das Potenzial einer osteoblastären Differenzierung.
Die Anwendung determinierter osteoblastärer Zellen hat den Vorteil, dass, so lange autologe Zellen verwendet werden, keine Immunabstoßung erfolgt und auch keine rechtlichen und ethischen Einschränkungen bestehen. Daher ist diese Zellart derzeit eine sehr vielversprechende bezüglich der knöchernen Regeneration. Zur Geweberegeneration ist allerdings eine ausreichende Anzahl von Zellen nötig, und somit ist man an die Kultivierbarkeit der Zellen gebunden. Inzwischen ist die Kultivierung dieser Zellen, welche unterschiedlichsten Ursprungs sein können, möglich [Meyer et al., 2005a]. Zunächst muss das Ursprungsgewebe (Knochen, Periost, Knochenmark) explantiert werden. Dann werden die Zellen mittels unterschiedlichen Techniken (mechanische Zersetzung, Enzymdigestion) aus dem Gewebe isoliert [Vacanti et al., 1993; Handschel et al., 2006]. Die isolierten Zellen können dann gemäß den Grundsätzen des Tissue Engineerings weiter proliferiert und differenziert werden.
Matrix
Knochen ist ein spezialisiertes Stützgewebe, das durch die Mineralisation von Osteoid, der nicht kalzifizierten extrazellulären Matrix ECM, entsteht. Die knöcherne Regeneration wird hauptsächlich durch Osteoblasten getragen, welche sich aus mesenchymalen Vorläuferzellen differenzieren. Diese Osteoblasten sezernieren die ECM, welche zu 90 Prozent aus Kollagen I besteht [Löffler, 2000], einem Marker für mesenchymale Stammzellen [Bilezikian et al., 1996]. In der Folge werden der ECM Kalzium- und Phosphationen zugeführt und es entstehen Hydroxylapatitkristalle (Geflechtknochen). Neben Kollagen I sind auch Osteopontin, Osteonectin und Osteocalzin sowie die Alkalischen Phosphatase Proteine, die im Verlauf der Knochenbildung sezerniert werden und typische Knochenmarker darstellen [Aubin et al., 1996; McKee et al., 1996; Termine et al., 1996; Zernik et al., 1990]. Es ist somit klar, dass an die optimale Matrix beziehungsweise das optimale Trägergerüst (Scaffold) für die Knochenneubildung viele Anforderungen gestellt werden. Es soll nicht toxisch, biokompatibel, biologisch abbaubar (unter Bildung nicht toxischer Abbauprodukte), nicht immunogen, einfach chirurgisch fixierbar sein, dabei gleichzeitig die individuelle interne und externe Knochengeometrie nachahmen, die Knochenbildung und das Zellattachment fördern sowie durch lokale Zellen umgebaut (remodelling) werden können [Harlan et al., 2002; Wiesmann et al., 2004]. Somit erscheint es offensichtlich, dass das optimale Trägergerüst derzeit nicht existent ist.
Scaffolds können in natürliche und künstliche Materialien unterteilt werden, wobei jede Klasse noch einmal in organische und anorganische Gruppen unterteilt wird. Somit werden synthetisch-organische (Polylactide, Polyglykoide), synthetischanorganische (Hydroxylapatit, Kalzium- Phosphat-Composite, Glaskeramiken), natürlich-organische (Kollagen, Fibrin, Hyaluronsäure) und natürlich-anorganische Materialien (Korallenhydroxylapatit) unterschieden. Die Zusammensetzung der Matrixkomponenten determiniert die Eigenschaften, wie Formsteifigkeit und Bruchfestigkeit, des Trägergerüstes [Wiesmann et al., 2004]. So werden die oben genannten Materialien in verschiedenen Konsistenzen (Gele, spongiöse Konstrukte, hoch komplexe Strukturen mit Poren und Kanälen) hergestellt [Griffith, 2002]. Neben dem langsamen Abbau und der Möglichkeit des Ersatzes durch Knochen, das Trägermaterial sollte möglichst ein geeignetes Substrat für die Enzyme der extrazellulären Matrix (ECM) darstellen, ist die Nachahmung der internen und externen Knochenmorphologie entscheidend für ein gutes Knochen- Scaffold [Langer et al., 1993]. Einflussgrößen, die den Nachahmungsgrad der Knochmorphologie beeinflussen sind unter anderen Dreidimensionalität, Materialoberfläche, Porengröße, Kanalausrichtung und Trabekelorientierung [Meyer et al., 2005b]. Diese physiko-chemischen Eigenschaften beeinflussen direkt die Zytokompatibilität (Zellattachment und Zellproliferation) des Trägermaterials. Da die Zellproliferation ein direktes Maß der Zytokompatibilität darstellt, untersuchte unsere Arbeitsgruppe die Proliferationsrate mittels CyQUANT®-Assay und Elektronenmikroskopie von ESC auf verschiedenen Trägermaterialien. Die Proliferation war am ausgeprägtesten auf bovinem ICBM (insoluble collagenous bone matrix) gefolgt von ß-Tricalciumphosphat multiporös (Cerasorb M®), ß-Tricalciumphosphat kleinporig (Cerasorb®), Copolymer aus Polylactat und Polyglycolsäure (PLA/PGA) sowie anorganischem bovinem Knochen (Bio Oss®) (Abbildung 3) [Handschel et al., 2008b]. Des Weiteren spielen die physiko-chemischen Eigenschaften eine entscheidende Rolle bei der Kompatibilität der Zellen auf einem Trägermaterial. ICBM besteht zu einem wesentlichen Anteil aus Kollagen I und zeigte in unseren Analysen die besten Voraussetzungen für die Zellproliferation. Diese Ergebnisse stehen im Einklang mit verschiedenen Studien, die nachwiesen, dass das Zellattachment von Osteoblasten von der Proteinbedeckung der Biomaterialoberfläche abhängt [Dennis et al., 1992; Meyer et al. 1998; Petrovic et al., 2006].
Des Weiteren konnten wir zeigen, dass das verwendete Trägermaterial Einfluss auf das Genexpressionsmuster der ESC hat [Handschel et al., 2008b]. Es ist allerdings noch unklar, wie diese Veränderung des Genexpressionsmusters durch die unterschiedlichen Biomaterialien den Differenzierungsprozess beeinflusst. Auch die Kräfte, denen ein Material oder Konstrukt in vivo ausgesetzt ist, beeinflussen die Resorbierbarkeit und damit die Materialeigenschaften [Handschel et al., 2002].
Derzeit herrscht eine Diskussion darüber, ob eher künstliche Scaffolds oder die natürliche Matrix verwendet werden sollten. Die Mikromassentechnologie ist ein Ansatz, um den Anteil an artifiziellen Scaffolds zu minimieren. Dabei werden Zellen nach der Proliferation von einer 2-D-Monolayer abgelöst und in spezielle, nicht adhäsive, Kulturkammern überführt. Dort bilden sich innerhalb von drei Tagen sphärische Formationen (Abbildung 4) [Handschel et al., 2007; Meyer et al. 2005a]. Diese Methode spiegelt eher die In-vivo-Bedingungen wider, da es sich hier im Gegensatz zu den Monolayerkulturen um eine dreidimensionale Kultur handelt. Es ist bekannt, dass die Verbindung zur ECM, zum Beispiel über Integrine, viele zelluläre Funktionen, wie Proliferation, Zelldifferenzierung, Zellmigration oder Apoptose, beeinflussen kann [Boudreau et al., 1999]. Bei dieser 3-D-Kulturform sezernieren die Zellen die ECM selbst, und die Zellen können in dem gewonnenen Konstrukt, welches frei von künstlichem Material ist, interagieren. DesWeiteren konnten in den Mikromassenkulturen höhere Proliferationsraten als in den Monolayerkulturen nachgewiesen werden. Mikromassenkulturen mit osteoblastenähnlichen Zellen zeigen eine schnellere und ausgeprägtere osteoblastäre Differenzierung als in 2-D-Kulturen [Gerber et al., 2001 und 2002]. Dass eine chondrogene Differenzierung in einer Mikromassenkultur möglich ist, ist bekannt [Naujoks et al. 2008], aber erst kürzlich konnte nachgewiesen werden, dass Mikromassenkulturen auch mit ESC möglich sind und mit ihnen knorpelähnliche Differenzierung möglich ist [Takana et al., 2004]. Zwischenzeitlich konnte unsere Arbeitsgruppe den Nachweis erbringen, dass eine osteogene Differenzierung von ESC in Mikromassen möglich ist [Handschel et al., 2008a].
Wachstumsfaktoren
Neben den Zellen und der Matrix sind spezifische Wachstumsfaktoren die dritte wichtige Säule des Tissue Engineering. Knochen ist ein lebendiges Gewebe, welches ständigen Umbauvorgängen unterliegt und sich somit ständig an die wechselnden Beanspruchungen anpasst. So ist bekannt, dass eine verminderte Belastung des Knochens zur Atrophie führt. Dies ist beispielsweise der Fall, wenn in einen zahnlosen Kiefer keine physiologischen Reize mehr eingeleitet werden und es somit zur Atrophie kommt. Diese biophysikalische Beeinflussbarkeit des Knochens, kann man sich im Tissue Engineering zunutze machen, indem man mechanische Reize zur Beeinflussung des Gewebes anwendet [Wiesmann et al., 2004]. Externe mechanische Kräfte haben direkten Einfluss auf die osteoblastäre Proliferation, die Zellorientierung und die Genaktivität. Brown et al. konnte beispielsweise nachweisen, dass eine zyklische Zugbeanspruchung zu einer Veränderung der Zellorientierung und der Genaktivität führt [Brown et al., 1998]. Osteoblasten sind sehr sensitiv auf mechanische Deformationen. Ist es möglich, die Kraftübertragung vom Scaffold auf den Osteoblast zu übertragen, führt dies zu einer erhöhten Osteoblastenproliferationsrate [Buckley et al., 1988; Meyer et al., 1999c; Neidlinger-Wilke et al., 1994] und zu einer Veränderung der Expression knochenspezifischer Proteine, wie alkalische Phosphatase, Osteopontin und Osteokalcin [Meyer et al., 1999c; Harter et al., 1995; Hillsley et al., 1994]. Der Mechanismus dieser Veränderungen ist noch nicht vollständig geklärt [Jones et al., 1995]. Neben der mechanischen Stimulation besteht die Möglichkeit der Stimulation mit piezoelektrischen Potenzialen, welche Osteoblasten über elektrische Felder beeinflussen [Hartig et al., 2000]. Eine elektrische Langzeitstimulation von Osteoblasten führt zu einer Veränderung des Genexpressionsmusters und zu einer die Knochenbildung fördernden erhöhten Synthese von extrazellulärer Matrix [Wiesmann et al., 2001]. Eine weitere Möglichkeit, die osteogene Differenzierung zu unterstützen, ist die biochemische Stimulation durch Zytokine und bioaktive Proteine [Schliephake et al., 2002]. Dabei scheint die gleichzeitige Anwendung von mehreren Faktoren die Knochenbildung zu unterstützen [Lieberman et al., 2002]. Da es entscheidend ist, dass die Faktoren in der angemessenen Dosis und zum richtigen Zeitpunkt an das Substrat gelangen, ist die Entwicklung entsprechender Trägermaterialien wichtig. Ein Reihe verschiedener Trägermaterialien (Kollagen Typ I, Hyaluronsäure) wurden bereits experimentell, aber auch in klinsichen Studien getestet [Bostrom et al., 1996; Gerhart et al., 1993; Radomsky et al., 1998 und 1999; Yasko et al., 1992]. Während der Knochenbildung werden viele verschieden biologisch aktive Moleküle exprimiert, wovon einige das Potenzial zur Förderung der Knochenregeneration besitzen [Schliephake et al., 2002; Terheyden et al., 2001]. Zu diesen Faktoren zählen Transforming Growth Faktor-ß (TGF-ß), Bone Morpheogenic Proteins (BMP), Fibroblast Growth Faktor (FGF), Plateletd-derived Growth Factor (PDGF) und Insulin-like Growth Factor (IGF). TGF-ß gehört zu der Familie von Proteinen, die eine Reihe von zellulären Aktivitäten, wie Wachstum, Differenzierung und Synthese extrazellulärer Matrix, regulieren. Zu der TGF-ß-Familie gehören ebenfalls die BMPs. 13 Moleküle wurden zwischenzeitlich identifiziert [Schmitt et al., 1999], wobei nach derzeitigem Wissensstand die BMPs-2, -4 und -7 durch Stimulierung der Differenzierung von mesenchymalen Zellen zu Osteoblasten eine wichtige Rolle bei der Knochenheilung übernehmen [Lieberman et al., 1998; Schmitt et al., 1999]. Durch die Entdeckung der Gensequenz der BMPs im Jahre 1988 ist es heutzutage möglich, mittels rekombinanter Gentechnologie verschiedene BMPs zu produzieren und sie auf verschieden Trägermaterialien aufzubringen [Wozney et al., 1988]. An Kaninchen mit einem Knochendefekt kritischer Größe konnte gezeigt werden, dass die Behandlung mit einem rhBMP-2-polylactid Scaffold, verglichen mit Kontrollen, zu besseren biomechanischen Ergebnissen mit höherer Radioopakizität führten [Wheeler et al., 1998]. Bioreaktoren scheinen ein Erfolg versprechendes Tool zu sein, um extendierte biologische Konstrukte in ausreichender Menge herzustellen [Depprich et al., 2008].
Zusammenfassung
Die Entwicklung des osteogenen Tissue Engineerings stellt einen Erfolg versprechenden Ansatz der regenerativen Medizin zur knöchernen Rekonstruktion dar. Die Forschung im Bereich der drei Säulen des Tissue Engineering hat noch viele Fragen zu klären. So muss geklärt werden, welche Zellen die optimalen Eigenschaften zur knöchernen Regeneration besitzen und durch welche Wachstumsfaktoren sich ihre knöcherne Regeneration steuern lässt. Insbesondere die Dosis und der Zeitpunkt der Applikation sowie die Applikationsart stellen noch große Herausforderungen dar. Des Weiteren ist es wichtig, ein Scaffold zu entwickeln, welches biokompatibel und biologisch abbaubar ist und gleichzeitig die biophysikalischen und biochemischen Anforderungen erfüllt, um eine schnelle und komplette Knochenregeneration zu gewährleisten. Auch, wenn noch längst nicht alle Probleme und Fragen gelöst sind, eröffnet das Tissue Engineering von Knochen vielversprechende Perspektiven für den Knochenersatz.
Dr. med. Christian NaujoksDr. med. Dr. med. dent. Rita DepprichPD Dr. med. Dr. med. dent. Jörg HandschelKlinik für Kiefer und Plastische Gesichtschirurgie,Westdeutsche Kieferklinik,Universitätsklinik Düsseldorf40225 DüsseldorfMoorenstraße 5
Christian.naujoks@med.uni-duesseldorf.deDepprich@med.uni-duesseldorf.deHandschel@med.uni-duesseldorf.de
")
