Parodontitis: Gene als Risikofaktor


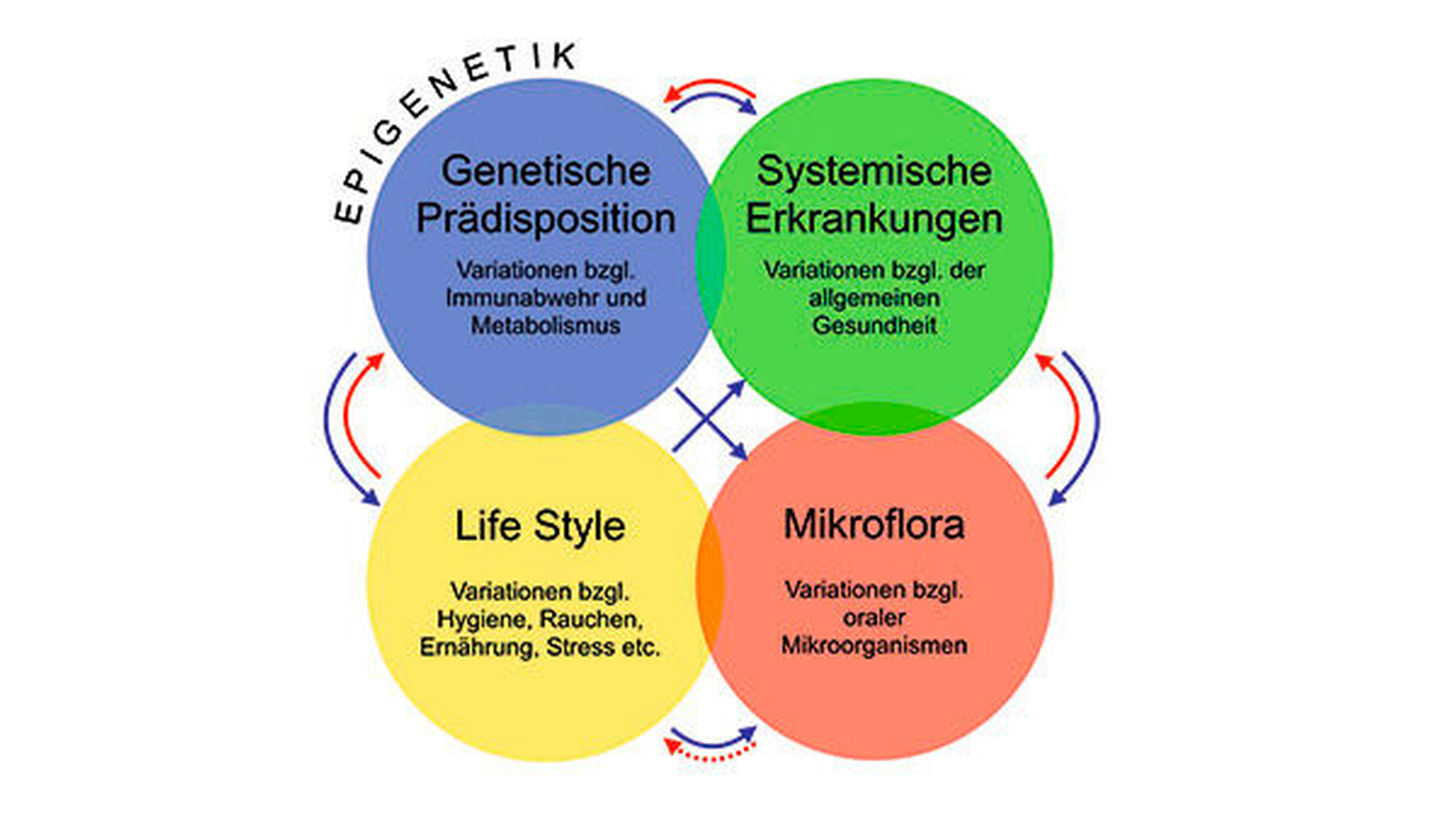

Die Parodontitis ist eine bakteriell induzierte entzündliche Erkrankung [Jepsen und Dommisch, 2014]. Weltweit wird die Prävalenz schwerer Parodontitis auf 10,5 bis 12 Prozent geschätzt, sie ist damit die sechsthäufigste Erkrankung der Menschheit [Kassebaum et al., 2014]. Die Parodontitis gilt als komplexe Erkrankung, da die individuelle Krankheitsanfälligkeit durch das Zusammenspiel zwischen dem oralen Mikrobiom, dem Immunsystem, Lifestyle-Faktoren, wie zum Beispiel Rauchen und Ernährung, und metabolischen Erkrankungen, wie Diabetes / metabolisches Syndrom bestimmt und durch Gene reguliert wird (Abbildung 1).
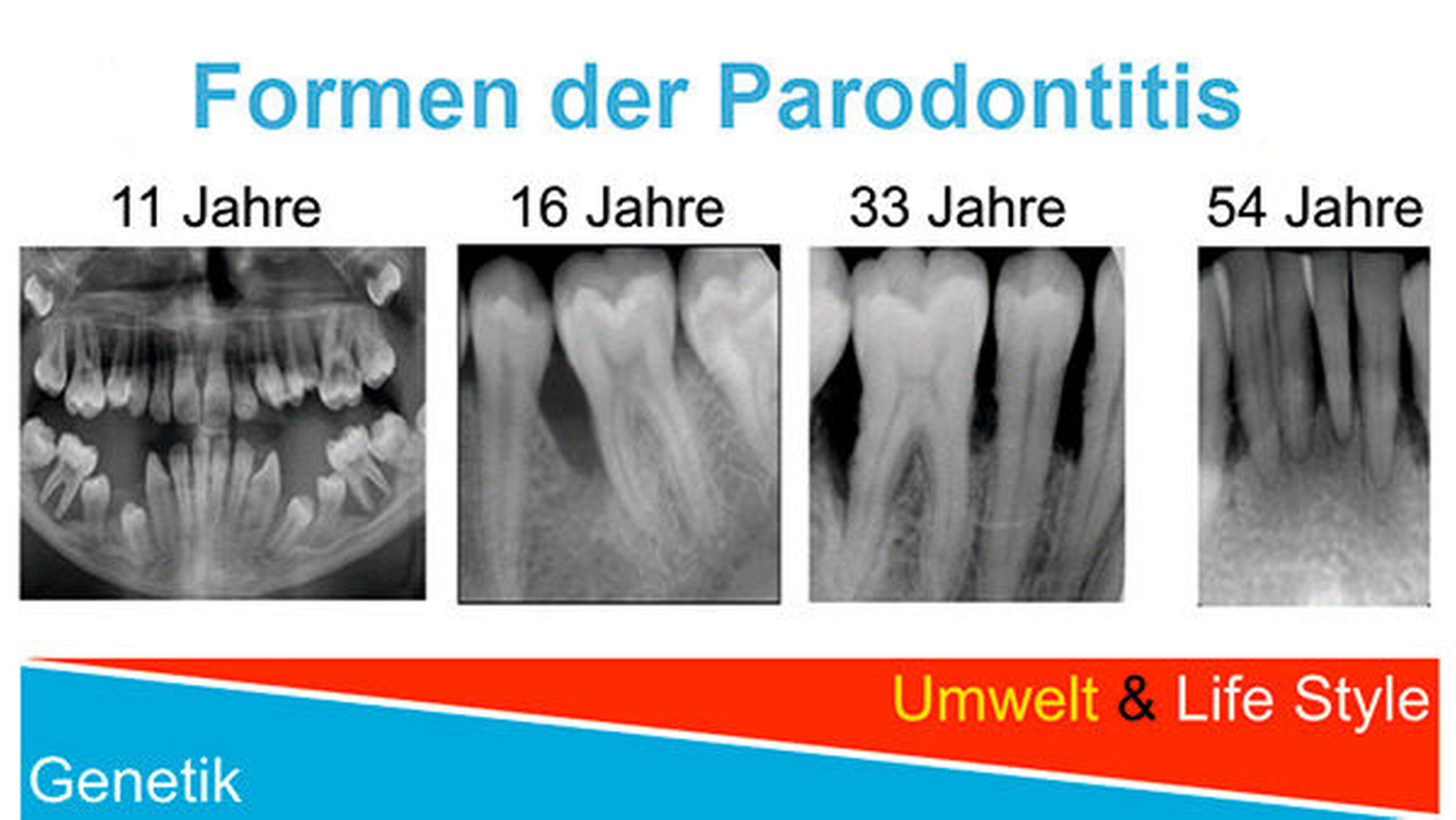
Nach ihrem klinischen Verlauf werden die seltenere aggressive (AgP) und die häufigere chronische Parodontitis (CP) unterschieden. Aufgrund der familiären Häufung, der raschen Progression und der frühen Manifestation der aggressiven Parodontitis (Abbildung 2) wird vermutet, dass genetische Faktoren für die individuelle Anfälligkeit an AgP zu erkranken eine besonders wichtige Rolle spielen. Im Gegensatz zur chronischen Parodontitis, bei der sich die negativen Effekte von Lifestyle- und Umweltfaktoren unter Umständen jahrzehntelang akkumulieren konnten, wird bei der aggressiven Parodontitis ein relativ stärkerer Anteil genetischer Einflussfaktoren vermutet (Abbildung 3). Dementsprechend wird in genetischen Studien zur Parodontitis in der Regel zunächst der Phänotyp der aggressiven Parodontitis untersucht. Genetische Forschung kann in besonderem Maß dazu beitragen, die Ursachen einer Erkrankung zu verstehen und zu erklären, warum sich Individuen in ihrer Krankheitsanfälligkeit unterscheiden, obwohl sie unter sehr ähnlichen Bedingungen leben.
Identifizierungsstrategien krankheitsrelevanter Gene
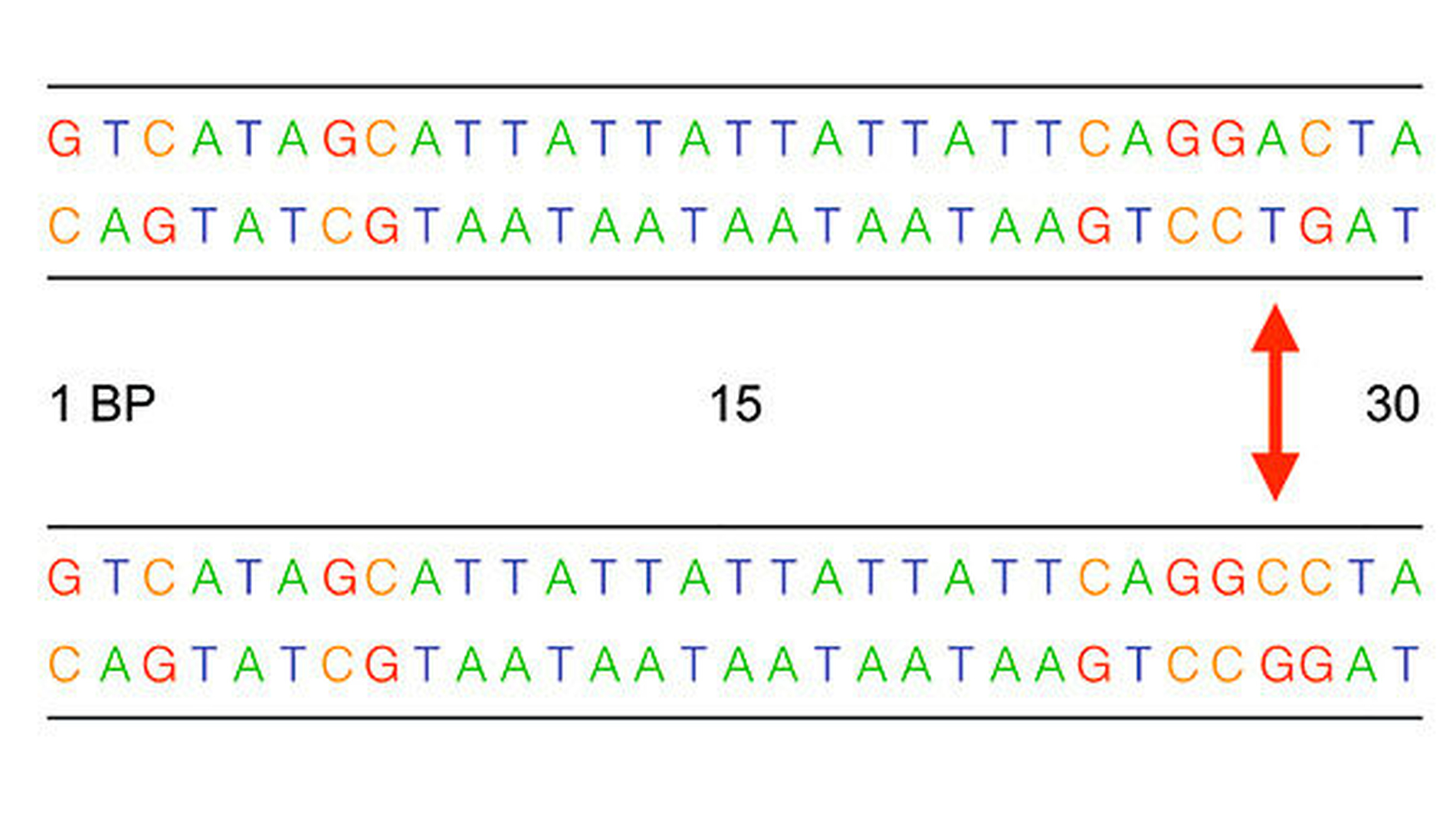
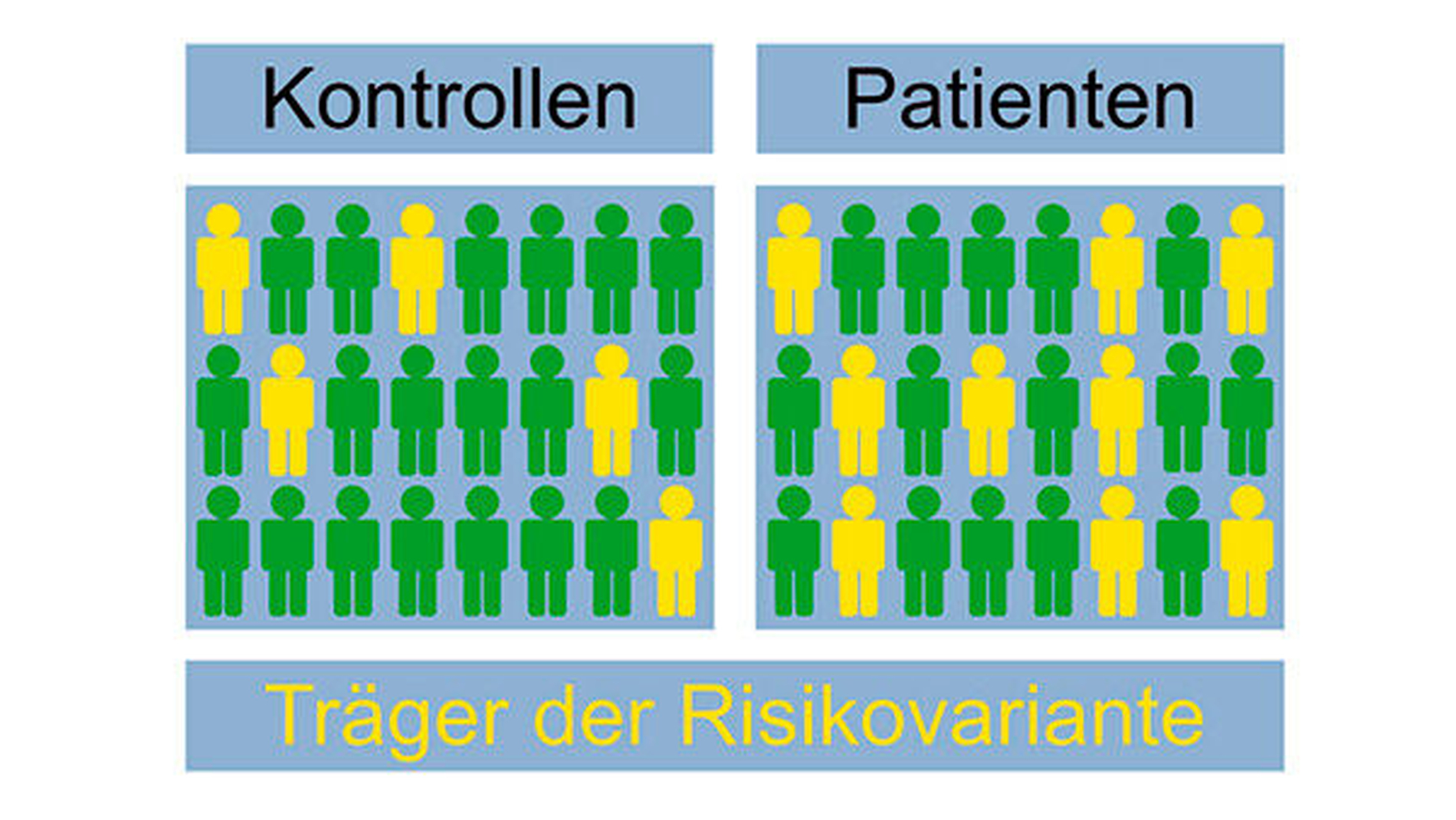
Im Mittelpunkt der genetischen Forschung zur Identifizierung krankheitsrelevanter Gene stand lange Zeit die Analyse sogenannter Einzelnukleotid-Polymorphismen (Single Nucleotide Polymorphism – SNP). Dabei handelt es sich um die Variation einzelner Basenpaare im DNA-Strang (Abbildung 4). Diese stellt die häufigste Form einer genetischen Variation dar. In Fall-Kontroll-Studien ausreichender Größe wird die Häufigkeit der Einzel-nukleotid-Polymorphismus(SNP)-Allele in zwei definierten Gruppen nicht verwandter Individuen verglichen: „Kontrollen“ ohne Erkrankung und „Fälle“ mit Erkrankung. Eine signifikant erhöhte Häufigkeit eines SNP-Allels beziehungsweise Genotyps in der Fall-Gruppe weist auf ein damit verbundenes erhöhtes Krankheitsrisiko hin (Abbildung 5). Allerdings muss diese statistische Assoziation immer in einer unabhängigen anderen Patientengruppe bestätigt werden.
Kandidatengen-Studien:Bis vor einigen Jahren war die Untersuchung von sogenannten Kandidatengenen, die nach dem aktuellen Verständnis der Krankheit und der jeweils verfügbaren Literatur ausgewählt wurden, die wichtigste Strategie für die Identifikation von Krankheitsgenen. Das besondere Kennzeichen von Kandidatengen-Studien ist die Erfordernis einer A-priori-Hypothese über die Beteiligung bestimmter Gene am Krankheitsrisiko sowie über die Anwesenheit bestimmter funktionaler Varianten innerhalb dieser Gene. Grundsätzlich gibt es zwei unterschiedliche Auswahlkriterien für ein Kandidatengen. Es kann beispielsweise interessant sein zu fragen, ob ein bestimmtes Gen innerhalb eines regulatorischen Signalwegs liegt, der für die Krankheit eine Bedeutung hat. Oder es gibt bereits funktionale Hinweise zu Varianten innerhalb eines Gens, die die Anfälligkeit für andere Krankheiten erhöhen.
Beide Ansätze geben Antworten auf die Frage, ob die ausgewählten Gene Varianten enthalten, die das Risiko für die untersuchte Krankheit erhöhen. Da jedoch die Hypothese für die Selektion eines bestimmten Gens vollständig vom gegenwärtigen Kenntnisstand der molekularbiologischen Mechanismen einer Krankheit abhängt, werden notwendigerweise Hunderte von Genen, die einen Einfluss auf die Krankheit haben können, nicht ausgewählt, da deren Funktion entweder unbekannt ist, oder weil diese in funktionellen Wirkketten liegen, die bis dahin noch nicht mit der Erkrankung in Verbindung gebracht wurden. Da die Ursachen vieler Erkrankungen oft sehr komplex und nur unvollständig verstanden sind, verwundert es nicht, dass die meisten der bisher veröffentlichten Krankheitsassoziationen der ausgewählten Gene nicht erfolgreich repliziert werden konnten.
Genomweite Assoziationsstudien (GWAS):Im Gegensatz zu Kandidatengen-Assoziationsstudien folgen sogenannte Genomweite Assoziationsstudien (GWAS) eher dem klassischen Ziel der Molekulargenetik, die Gene oder Signalwege direkt zu identifizieren, die das jeweilige Krankheitsrisiko beeinflussen. In diesem Studientyp wird eine sehr hohe Anzahl DNA-Sequenzvarianten (SNPs), die über das gesamte Genom verteilt sind und den größten Teil der genetischen Information beschreiben, in den Patienten und gesunden Kontrollen untersucht. Daher bilden GWAS einen hypothesenfreien und damit unvoreingenommenen Zugang. Da das Design von GWAS die genaue Kenntnis der Sequenzunterschiede im Humangenom voraussetzt und auch erst die technischen Voraussetzungen vorhanden sein mussten, um die Identität von oftmals über einer Million untersuchter genetischer Varianten pro Individuum zu bestimmen, werden GWAS im Wesentlichen erst nach Veröffentlichung der vollständigen annotierten Sequenz des letzten Humanchromosoms im Jahr 2006 durchgeführt [Gregory et al., 2006].
Beginnend im Jahr 2007 [WTCCC, 2007] lag einer der wesentlichsten Schwerpunkte humangenetischer Forschung in der Aufklärung der Bedeutung von DNA-Sequenzvarianten für komplexe Erkrankungen. Und seither wurden über 10 000 krankheitsasso- ziierte Varianten veröffentlicht [Welter et al., 2014]. Für viele Erkrankungen ist mit Meta-Analysen, die Zehntausende von Patienten und Fällen einschlossen, die systematische genomweite Suche nach krankheitsverursachenden Genvarianten mit Hunderten replizierter Assoziationen zu einem voraussicht- lichen Abschluss gekommen [Schreiber et al., 2014].
Die zuletzt genannten Zahlen zur Größe der verwendeten klinischen Analysepopulationen verdeutlichen, dass für Assoziationsstudien aus statistischen Gründen sehr hohe Fallzahlen benötigt werden, die in der Regel allein für die erste explorative Studie weit über Tausende von Patienten und die doppelte Anzahl gesunder Kontrollen benötigten. Die notwendige Replikation der Befunde erfordert eine weitere unabhängige Stichprobe von mindestens der gleichen Größe [Schäfer et al., 2011a].
Da die nötige hohe Anzahl gut diagnostizierter Patienten, die in diese Studien ein- geschlossen werden müssen, in der Zahnmedizin bis in die junge Vergangenheit nicht vorhanden war, täuscht die hohe Zahl an Veröffentlichungen von Kandidatengen-Assoziationsstudien in der zahnmedizinisch-wissenschaftlichen Literatur darüber hinweg, dass bislang nur sehr wenige Studien mit ausreichender statistischer Teststärke durchgeführt wurden [Vaithilingam et al., 2014]. Im Folgenden werden diejenigen Gene vorgestellt, die in Fall-Kontroll-Studien mit ausreichender Teststärke identifiziert und repliziert oder durch verschiedene Ansätze mehrfach unabhängig bestätigt werden konnten.
Genetische Risikofaktoren der Parodontitis
ANRIL (Antisense RNA in the Inc locus):Das Gen ANRIL wurde im ersten GWAS der Atherosklerose identifiziert [WTCCC, 2007] und ist in der Medizin weltweit als der wichtigste und am besten replizierte genetische Risikofaktor für den Herzinfarkt bekannt [Deloukas et al., 2013]. Weniger bekannt ist, dass ANRIL auch als das am besten replizierte Risikogen für die Parodontitis gilt [Schäfer et al., 2009; Ernst et al., 2010; Schäfer et al., 2011b; Schäfer et al., 2013; Bochenek et al., 2013]. ANRIL ist durch den STAT1(Signal Transducer and Transcription 1)-Signalweg reguliert, der die Entzündungsantwort nach Stimulierung mit dem ent-zündungsfördernden Zytokin Interferon-γ steuert [Harismendy et al., 2011].
Neue Studien weisen darauf hin, dass ANRIL ebenfalls durch den TGF-β-Signalweg reguliert ist [Schäfer et al., 2015]. Interessanterweise enthält das Gen ANRIL nicht die Erbinformation für ein Protein, sondern codiert eine lange Ribonukleinsäurekette (RNA-Transkript). Diese Moleküle haben eine wichtige Funktion in der Regulation der Aktivität anderer Gene [Mercer et al., 2009]. Entsprechend konnte eine Regulation der ANRIL benachbarten Gene CDKN2A und -2B (Cykline Depending Kinase 2A und -2B) nachgewiesen werden, die eine wichtige Funktion in der Regulation der Zellteilung haben [Yap et al., 2010].
Es ist aber nicht sicher, ob diese Funktion für das erhöhte Risiko für Parodontitis und Atherosklerose verantwortlich ist, denn lange nicht-protein-codierende RNA-Moleküle wie ANRIL haben oft auch regulatorische Effekte auf im Genom weit entfernt liegende Gene. So konnte eine genomweite Untersuchung derartiger trans-Effekte nachweisen, dass eine induzierte Verminderung der ANRIL-Moleküle einen direkten Effekt auf die biologische Aktivität der Gene ADIPOR1 (Adiponectin Receptor 1), C11ORF10 (chr.11 open reading frame 10) und VAMP3 (Vesicle-associated membrane protein 3) hat [Bochenek et al., 2013]. ADIPOR1 ist ein Rezeptor für das Proteinhormon Adiponectin und vermittelt durch dessen Bindung den oxidativen Abbau von Fettsäuren und die Regulation der Glucoseaufnahme.
Im weiteren Sinn spielt Adiponectin damit eine wichtige Rolle in der Unterdrückung metabolischer Störungen, die zu Adipositas, Atherosklerose und Typ-2-Diabetes führen können. Das bisher nicht näher charakterisierte Transmembranprotein C11ORF10 liegt innerhalb des FADS(Fatty Acid Desaturase)-Genclusters. Bei diesem handelt es sich um eine Gruppe von Genen, die eine wichtige Rolle in der metabolischen Regulation des Verhältnisses gesättigter zu ungesättigten Fettsäuren spielen.
Entsprechend zeigten GWAS Assoziationen genetischer Varianten innerhalb von C11ORF10 und der FADS- Gene mit Typ-2-Diabetes [Dupuis et al., 2010], dem Metabolischen Syndrom [Lemaitre et al., 2011; Zabaneh und Balding, 2010] sowie der chronisch entzündlichen Darmerkrankung Morbus Crohn [Franke et al., 2010]. Eine nachfolgende genetische Untersuchung zeigte auch eine schwache, aber nominal signifikante Assoziation dieser Varianten mit AgP [Bochenek et al., 2013]. Zusammenfassend hat ANRIL vermutlich einen Einfluss auf die Regulation des Zell- zyklus und des Metabolismus von Makronährstoffen.
VAMP3:Bevor die ätiologische Bedeutung von VAMP3 durch den funktionalen Zusammenhang mit ANRIL gezeigt wurde, war VAMP3 bereits durch eine andere genomweite Studie ins Blickfeld der parodontologischen Forschung gerückt [Divaris et al., 2012]. In dieser Studie wurden Assoziationen bestimmter oraler Bakterien mit Nukleotidvarianten auf genomweiter Ebene untersucht. Dabei wurde gezeigt, dass die chromosomale Region von VAMP3 hochsignifikant mit dem erhöhten Vorkommen von Pathogenen des sogenannten „orange complex“ assoziiert ist. Pathogene, die zu diesem bakteriellen Komplex zusammenge fasst werden (wie Fusobacterium nucleatum), sind Bakterien, die aufgrund ihres Stoffwechsels in der Mundhöhle eine ökologische Nische für anaerobe Bakterien wie zum Beispiel Porphyromonas gingivalis schaffen die als für die Destruktion des Parodonts ursächliche Keystone-Pathogene angesehen werden [Hajishengallis, 2014].
Diese GWAS hatte gezeigt, dass genetische Varianten innerhalb von VAMP3 – und bis in das oberhalb von VAMP3 liegende Gen CAMTA1 (calmodulin binding transcription activator 1) hineinreichend – mit einem erhöhten Vorkommen von Bakterien dieses Komplexes assoziiert sind. Eine genaue Feinkartierung dieser Region mit Patienten der AgP iden- tifizierte schließlich eine seltene Variante in CAMTA1 als mit AgP assoziiert [Bochenek et al., 2013]. Entsprechend der Assoziation mit ANRIL wurde diese Assoziation dann in einer Stichprobe von 21 000 Herzinfarktpatienten und 44 000 gesunden Kontrollen als auch mit dem Herzinfarkt assoziiert bestätigt [Bochenek et al., 2013]. Die genaue biologische Funktion des einen Transkriptionsfaktor codierenden CAMTA1-Gens ist derzeit noch unbekannt, Assoziationen zu den seltenen Erkrankungen „inflammatorisches arteriovenöses Hämangiom“ (eine vaskuläre Fehlbildung mit sekundärer Entzündung) und „zerebelläre Ataxie“ (Störungen der Bewegungskoordination durch pathologische Veränderungen im Kleinhirn ausgelöst) wurden für CAMTA1 jedoch beschrieben [Thevenon et al., 2012]. Im Gegensatz dazu hat VAMP3 verschiedene, gut untersuchte Funktionen in der TNF-α-vermittelten Phagozytose durch Makrophagen [Murray et al., 2005] und interessanterweise auch in der Regulation der zellulären Verfügbarkeit von Glukose [Schwenk et al., 2008].
Die möglichen funktionalen Effekte der AgP-, Atherosklerose- und Bakterien-assoziierten Varianten sind allerdings bislang noch un-bekannt. Denkbar ist aber, dass diese die Regulation der Aktivität des VAMP3-Gens beeinflussen. Zusammenfassend könnten mit VAMP3 assoziierte Risikovarianten auf den Zuckerstoffwechsel sowie auf immunologische Faktoren Einfluss nehmen. PLG (Plasminogen): Zusätzlich zu den Risikovarianten in ANRIL und CAMTA1 zeigt auch das Gen PLG hochsignifikante Krankheitsassoziationen mit AgP [Schäfer et al., 2015] und Atherosklerose [Deloukas et al., 2013] und gilt als der dritte derzeit bekannte gemeinsame Risikofaktor beider Krankheiten. Plasminogen ist der inaktive Vorläufer von Plasmin und wird an den Zellmembranen zu Plasmin prozessiert. Eine wichtige Aufgabe des Plasmin ist die Fibrinspaltung bei der Auflösung von Blutgerinnseln (Thromben). Plasmin hat aber auch die Fähigkeit, die Produktion inflammatorischer Zytokine zu stimulieren.
Die Identifikation von PAI-1 (Plasminogen activator inhibitor 1) als Inhibitor des PLG-Plasmin-Systems, das in viszeralem Fett stark exprimiert ist, weist auf die Verbindung zu Adipositas hin [Shimomura et al., 1996]. Das PLG-Plasmin-System hat durch den Plasmin-vermittelten Abbau von Gewebebarrieren aber auch für die Zellmigration eine wichtige Bedeutung. Interessanterweise können verschiedene pathogene Bakterien humanes PLG an bakteriellen Rezeptoren binden und durch diese Bindung proteolytische Eigenschaften erwerben. Beispielsweise ist Porphyromonas gingivalis in der Lage, die Expression humanen PLGs zu aktivieren und humane Plasmininhibitoren zu hemmen, wodurch eine vom Wirt nicht mehr kontrollierte proteolytische Plasminaktivität hervorgerufen wird [Boyle und Lottenberg, 1997; Grenier, 1996; Lahteenmaki et al., 2001]. Es bedarf jedoch funktionaler Studien, um die Mechanismen zu identifizieren, durch die die assoziierten Varianten das Risiko sowohl für die AgP als auch für die Atherosklerose vermitteln, und um zu klären, ob die jeweiligen Mechanismen voneinander abhängig sind.
NPY (Neuropeptide Y):Das Gen NPY rückte als mögliches Risikogen der Parodontitis zum ersten Mal durch eine GWAS zur schweren CP ins Blickfeld der Forschung [Divaris et al., 2013]. In dieser Studie wurden Varianten stromabwärts der proteincodierenden Bereiche von NPY als hochsignifikant mit schwerer CP assoziiert gefunden. Eine weitere GWAS, in der die Interaktionen des männlichen und des weiblichen Geschlechts mit Nukleotidvarianten in Patienten der AgP untersucht wurde, fand die statistisch aussagekräftigsten Effekte dieser genomweiten Studie stromaufwärts von NPY [Freytag-Wolf et al., 2014]. Geschlechtsspezifische Effekte von NPY wurden bereits in Versuchen mit Mäusen und mit Ratten beschrieben [Painsipp et al., 2011; Forbes et al., 2012]. So zeigten Mauslinien mit einem Funktionsverlust von NPY in Verhaltenstests unterschiedliche Stress-reaktionen in den verschiedenen Geschlechtern.
Des Weiteren zeigten diese Linien in Verhaltenstests unterschiedliche Reaktionen zwischen den Geschlechtern in Bezug auf Nahrungsaufnahme und Passage durch den Gastrointestinaltrakt. In diesem Zusammenhang ist ein weiterer interessanter Aspekt, dass NPY in Menschen eine Appetit-stimulierende Wirkung hat, die durch das Proteohormon Leptin unterdrückt wird. Eine weitere funktionale Studie, die eine Verbindung von NPY mit Parodontitis herstellte, zeigte höhere Konzentrationen von NPY in der Sulkus- flüssigkeit bei gesunder Gingiva im Vergleich mit einer durch eine Parodontitis entzündeten Gingiva [Lundy et al., 2009]. NPY ist das häufigste Neuropeptid des Knochens und es hat eine Funktion in der Erhaltung der Balance zwischen Knochenbildung und -resorption. Außerdem wird vermutet, dass NPY immunmodulatorische Effekte ausübt, indem es das Verhältnis der proentzündlichen T1-Helferzellen (Th1-Zellen) zu Th2-Zellen beeinflusst. Die Bindung von NPY an den NPY1-Rezeptor fördert vermutlich die anti-entzündliche Th2-Antwort. Aus diesen Gründen wird für NPY eine Bedeutung in der Koordination der Entzündungsreaktion und des Knochenmetabolismus angenommen.
GLT6D1 (glycosyltransferase 6 domain containing 1):
Das Gen GLT6D1 ist das bislang einzige bekannte Risikogen der Parodontitis mit einer Krankheitsassoziation von genomweiter Signifikanz und wurde entsprechend in der ersten GWAS zur AgP identifiziert [Schäfer et al., 2010]. Es ist derzeit auch das am besten replizierte AgP-spezifische Risikogen mit ätiologischer Bedeutung sowohl für die europäische als auch für die afrikanische Bevölkerung [Hashim et al., 2015]. Aufgrund von Sequenzhomologien wurde eine Hexose-Transferase-Aktivität annotiert (Gene Ontolgoy Consortium), doch die In-vivo-Funktion von GLT6D1, deren Spezifität und Substrate sind bisher vollkommen unbekannt.
Glycosylierungen durch Glycosyltransferasen haben eine wesentliche Bedeutung für die Funktion des Immunsystems, unter anderem in der Regulation der Signalwirkung der Rezeptoren von T-Zellen, B-Zellen, der Immunzelldifferenzierung und der Immunzellhomöostase sowie der Pathogenerkennung [Marth und Grewal, 2008; Rabinovich und Croci, 2012; van Kooyk et al., 2013]. Interessanterweise ist das wichtigste, derzeit bekannte Paralog von GLT6D1 die Glycosyltransferase GBGT1 (Globoside Alpha-1,3-N- Acetylgalactosaminyltransferase 1), die eine Rolle in der Synthese von Forssman-Glyco- lipiden erfüllt. Diese Glycolipide bilden zelluläre Bindestellen für Pathogene und können den Tropismus des Wirtes gegenüber Mikroorganismen determinieren [Xu et al., 1999]. Eine Funktion von GLT6D1 in der Determinierung des Wirtstropismus wird daher vermutet.
Ausblick
Die im Vergleich zu anderen komplexen Erkrankungen wenigen bisher durchgeführten, großen systematischen Studien zur Identifikation der genetischen Risikofaktoren der Parodontitis identifizierten eine begrenzte Zahl von Risikogenen, die eindeutig repliziert oder durch andere Ansätze unabhängig bestätigt werden konnten. Interessanterweise stehen diese Gene in einem Zusammenhang mit Mechanismen, die das Immunsystem und den Fett- und Kohlenhydrat-metabolismus verbinden. Diese Befunde weisen möglicherweise darauf hin, dass Störungen dieser Funktionen durch genetische Faktoren ein gemeinsames Merkmal der verschiedenen Phänotypen der Parodontitis sind. Die zum gegenwärtigen Zeitpunkt identifizierten Risikovarianten können nur einen geringen Anteil der Erblichkeit der Parodontitis erklären. Die Identifikation des vollständigen Spektrums krankheitsrelevanter Risikoallele kann jedoch nur gelingen, wenn die Erbinformation von Tausenden Patienten und gesunder Kontrollen systematisch untersucht wird.
Obwohl dazu erst seit ein paar Jahren die technischen Möglichkeiten bestehen, konnte bereits für andere komplexe Entzündungserkrankungen, für die zum Teil Hunderte replizierter Risikogene identifiziert wurden, das hohe Potenzial derartiger Studien gezeigt werden. So gilt beispielsweise für chronisch entzündliche Darmerkrankungen die Suche nach krankheitsverursachenden Genvarianten bereits als abgeschlossen. Derartige Forschungsprozesse lassen sich jedoch nur in großen Konsortien bewältigen, die international organisiert werden. Hier ist Deutschland in der genetischen Forschung zur Parodontitis international deutlich sichtbar, da die Deutsche Forschungsgemeinschaft (DFG) über die letzten Jahre diese Bemühungen weitsichtig und gezielt gefördert hat und fördert.
Derzeitige Implikationen für die Zahnarztpraxis
Die aufgrund der Qualität der Studien als validiert geltenden Risikogene weisen auf die Bedeutung der Interaktionen des metabolischen Systems mit dem Immunsystem in der Ätiopathogenese der Parodontitis hin. Dieser Interaktion liegen vermutlich viele der Komplikationen der Parodontitis und mit ihr assoziierter Komorbiditäten zugrunde – und es bietet sich möglicherweise in der Zukunft eine Vielzahl therapeutischer Möglichkeiten an. Allerdings befindet sich die Forschung hier noch in einer frühen Phase. Bevor Erkenntnisse aus der Genetik in die klinische Praxis übersetzt werden können, müssen zuvor noch grundsätzliche Fragen beantwortet werden.
So ist derzeit unbekannt, über welche molekularen Wirkketten Störungen des Fettstoffwechsels zur Entstehung der Parodontitis beitragen oder in welchem Ausmaß Diabetes, Adipositas und Entzündungsreaktionen parallel oder sich gegenseitig fördernd ausgelöst werden. Dennoch bestehen Erwartungen – durch die großen Fortschritte der Humangenetik in der medizinischen Forschung durchaus berechtigt – für Verbesserungen in Prävention, Frühdiagnose und Therapie. Diese Erwartungen liegen auch in der sogenannten „personalisierten Medizin“, die unter anderem auf der Feststellung der individuellen genetischen Profile der Patienten und den davon abzuleitenden Konsequenzen für eine „maßgeschneiderte“ Behandlung beruht. Gegenwärtig sind für die Parodontitis die komplexen Interaktionen zwischen genetischen Faktoren und Faktoren des Lebensstils, der Umwelt und des Alters aber noch unzureichend verstanden.
Die gewonnenen Ergebnisse der wenigen großen und systematischen genetischen Studien weisen auf eine hohe Heterogenität der chronischen Parodontitis hin, die durch genetische Anteile, aber auch stark durch die negativen additiven Effekte des Lebensstils, der Umwelt und altersbezogener Faktoren sowie deren Interaktionen geschaffen wird. Aufgrund der Komplexität dieser verschiedenen Faktoren kann grundsätzlich nicht erwartet werden, dass die Testung der Trägerschaft bestimmter häufiger DNA-Sequenzvarianten einzelner Gene es erlaubt, Patienten für eine frühzeitige Diagnose, eine indi vidualisierte Prävention und eine verbesserte Therapie zu stratifizieren [Ioannidis und Trikalinos, 2007; Ioannidis et al., 2008; Janssens et al., 2008]. Dies sind jedoch Hoffnungen, die
oftmals auch aus kommerziellen Interessen
geweckt werden. In der gegenwärtigen Situation, in der, wie eingangs beschrieben, die meisten veröffentlichten Assoziationen zum Risiko der Parodontitis wissenschaftlich nur unzureichend belegt sind, ist die Auswahl genetischer Varianten aus diesen Studien zur Erstellung eines individuellen Risikoprofils nicht zielführend, insbesondere wenn die Varianten nur moderate Assoziationen zeigen und eine funktionale Charakterisierung der angenommenen Effekte fehlt.
In diesem Zusammenhang wurden gegen die derzeit verfügbaren genetischen Tests von der Europäischen und von der Amerikanischen Gesellschaft für Humangenetik schwere Bedenken formuliert [Janssens et al., 2008; Patch et al., 2009]: Ein persönliches Risikoprofil muss, um klinische Bedeutung zu haben, sämtliche für die Ätiologie der Krankheit bedeutsamen Faktoren einschließen, die über die Trägerschaft einzelner Risikovarianten in bestimmten Genen hinausgehen. Ein derartiges Profil zu erstellen, würde das vollständige Wissen über alle Risikovarianten erfordern, sowie über ihre Interaktionen untereinander. Diese Interaktionen sind derzeit jedoch noch weniger gut verstanden. Im Gegensatz dazu sind für eine Marktzulassung genetischer Tests meist jedoch nur Belege für die analytische Validität des Tests erforderlich. Wird der jeweilige Genotyp zuverlässig identifiziert, gilt die im Test verwendete Methode als analytisch korrekt. Wenn dieser Genotyp jedoch nichts mit der untersuchten Krankheit zu tun hat, besitzt der Test keine klinische Validität und ist für den Patienten wertlos [Patch et al., 2009].
Zusammenfassung
Zusammenfassend ist festzustellen, dass genetische Risikoprofile zukünftig zwar das Potenzial haben können, die Effektivität und Effizienz präventiver Interventionen individuell zu verbessern. Derzeit sind aber die wissenschaftlichen Belege für die meisten der veröffentlichten Risikovarianten unzureichend. Die wenigen als validiert geltenden und in diesem Beitrag vorgestellten Risikogene und ihre entsprechenden assoziierten Varianten verleihen eine zu geringe Vorhersagegenauigkeit für eine dem Patienten gewinnbringende Anwendung in einem persönlichen genetischen Risikoprofil. Dennoch liefern die als validiert geltenden Risikogene der Parodontitis bereits wichtige Erkenntnisse für das Verständnis der Ätio- pathogenese der Parodontitis, indem sie sowohl auf eine ausgeprägte gemeinsame genetische Basis der aggressiven und der chronischen Parodontitis als auch auf die große Bedeutung der Interaktion des Fett- und Glukosemetabolismus mit dem Immunsystem für die Parodontitis hinweisen.
PD Dr. rer. nat. Arne Schäfer
Prof. Dr. med. dent. Henrik Dommisch
Abteilung für Parodontologie und Synoptische Zahnmedizin, Charité, Universitätsmedizin Charité – Universitätsmedizin Berlin
Aßmannshauser Str. 4-6, 14197 Berlin
und Dept. of Oral Health Sciences, University of Washington, Seattle, WA, USA
<link url="arne.schaefer@charite.de" target="new-window" url-fragment="" seo-title="" follow="follow">arne.schaefer@charite.de<link url="henrik.dommisch@charite.de" target="new-window" url-fragment="" seo-title="" follow="follow">henrik.dommisch@charite.de
Prof. Dr. med. dent. Dr. med. Søren Jepsen,
M.S.Poliklinik für Parodontologie, Zahnerhaltung, Präventive Zahnheilkunde
Universitätsklinikum Bonn
Welschnonnenstr. 17, 53111 Bonn
<link url="jepsen@uni-bonn.de" target="new-window" url-fragment="" seo-title="" follow="follow">jepsen@uni-bonn.de

